Astrocytes are considered the natural caretaker cells of the brain and are the most abundant cell type in the human brain 1,2. Astrocytes provide metabolic and trophic support to neurons and they modulate synaptic activity 3. After brain injury, astrocytes play pivotal roles in restoring ion balance, controlling edema, and removing toxic neurotransmitters 4-6. These neuroprotective activities are highly energy-dependent and require astrocyte mitochondria 7,8. Notably, neurons are permanently injured after ischemia only if astrocyte mitochondrial function fails 9,10. Inhibition of astrocyte mitochondria increases cell swelling and induces cell death11. Excitotoxicity, due to high extracellular glutamate, is also principally controlled by astrocytes and requires membrane polarization – the energy-dependent maintenance of the Na+ ion gradient12-14.
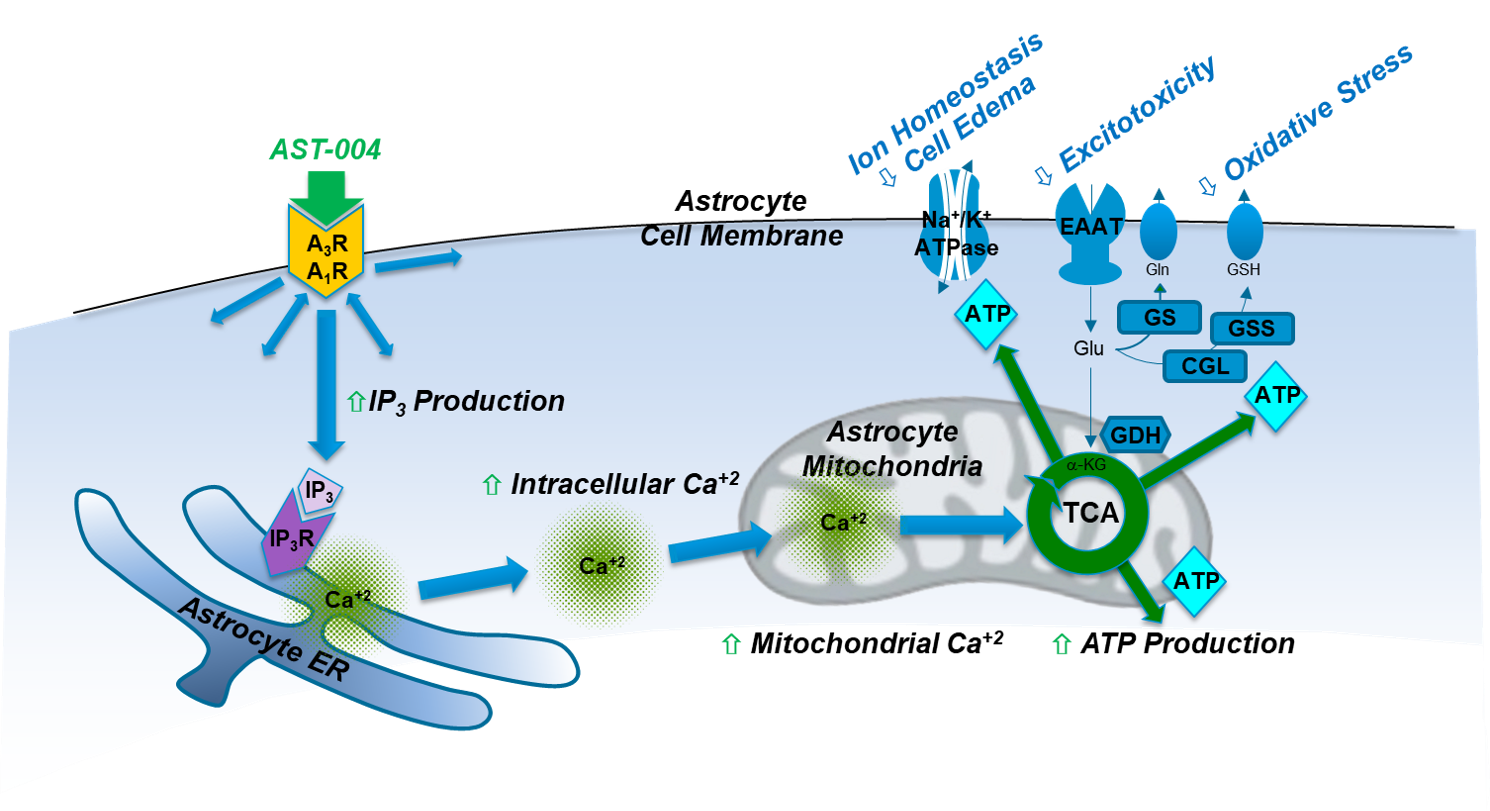
Adenosine is a ubiquitous endogenous compound that plays critical roles in nearly all aspects of cellular physiology throughout the body. Under normal physiological conditions, adenosine concentrations in the body are maintained at low nanomolar concentrations. However, under conditions of oxidative stress and trauma, dramatic increases in adenosine concentrations can be observed, leading researchers to classify the compound not as a hormone, but as a “retaliatory metabolite” with critical roles in maintaining cellular function. Extracellular increases in adenosine activate the adenosine A1 and A3 receptors on glial cells. On astrocytes, this produces inositol (1,4,5) triphosphate (IP3) that triggers intracellular Ca2+ release from thapsigargin-sensitive stores in the endoplasmic reticulum (ER), 15-18. During astrocyte activation, mitochondria, in turn, sequester some of the released Ca2+ and utilize it to activate Ca2+ sensitive dehydrogenases in the tricarboxylic citric acid cycle (TCA). This signaling cascade thereby increases oxidative phosphorylation and ultimately, significant production of ATP19-21.
Our results demonstrate that in multiple preclinical models of stroke and blunt trauma TBI, treatment with AST-004 significantly reduces early brain damage 17,18,22-26. These agonists selectively enhance mitochondrial ATP production in astrocytes by stimulating IP3-mediated Ca2+ release, which, in turn, enhance their energy-dependent neuroprotective functions. Pharmacological and genetic manipulation show that receptor activation not only increases neuronal and astrocyte survival but also partially reverses neuronal and glial damage. Our data also show that the protective pathway activated by our therapeutic candidates is conserved in ex vivo human brain tissue experiments (unpublished), emphasizing the potential for this novel approach to be an effective therapy for human brain injuries.